





All’interno del progetto AGER 4F è stato affrontato il problema della sostituzione della FM con VM, PM ed IM, individuando sperimentalmente la necessità di bilanciamento aminoacidico.
A titolo di esempio, come si vede dalla figura 1, una sostituzione di FM con farina di soia comporta l’apporto degli stessi aminoacidi (AA) contenuti nella proteina di soia (SP).
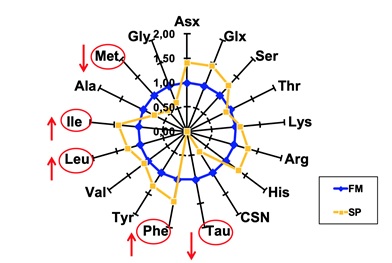
Fig. 1 – Comparazione della presenza di aminoacidi nella proteina di soia (SP) in giallo,
rispetto alla farina di pesce (FM) in blu, normalizzata uguale ad 1
A fronte di un aumento nella SP degli AA ramificati quali leucina ed isoleucina, quindi fenilalanina, istidina, arginina, serina, acido aspartico, acido glutammico, si assiste ad una carenza di aminoacidi essenziali, quali i solforati cisteina e metionina, ed una completa assenza dell’aminoacido funzionale taurina.
La metionina (Met) si può integrare con addizioni dell’aminoacido di sintesi oppure di altre farine proteiche che lo contengano, ripristinando la stessa quantità presente nella FM, ottenendo la comparazione riportata in fig. 2, dove comunque risulta evidente l’assenza totale di taurina.
L’equivalenza nella presenza di metionina nelle due sorgenti proteiche, dopo la dovuta integrazione, non significa comunque che l’aggiunta di metionina sia completamente efficace, ovvero abbia la stessa biodisponibilità
degli altri AA legati alle proteine.
Infatti, si presenta un problema di sincronismo, ossia la metionina introdotta in forma di cristalli, risulta velocemente disponibile, arrivando al fegato prima degli aminoacidi proteici, non potendo pertanto contribuire alla funzione proteinogenica. Gli aminoacidi legati alle proteine, giungendo al fegato in ritardo, si troverebbero comunque in carenza di metionina.
Infatti, si presenta un problema di sincronismo, ossia la metionina introdotta in forma di cristalli, risulta velocemente disponibile, arrivando al fegato prima degli aminoacidi proteici, non potendo pertanto contribuire alla funzione proteinogenica. Gli aminoacidi legati alle proteine, giungendo al fegato in ritardo, si troverebbero comunque in carenza di metionina.
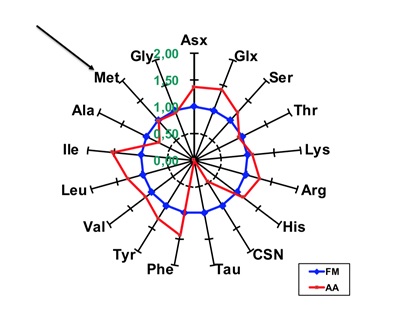
Fig. 2 - Comparazione di spettri aminoacidici come in Fig. 1,
dopo addizione di Met alla farina di soia.
Simile problema, sebbene a carico di diversi AA, lo incontriamo quando la FM viene sostituita con altre farine proteiche, quali le farine di insetti e di avicoli. Per alcuni aminoacidi essenziali si rende quindi necessaria una integrazione mirata a coprire le carenze presenti in ciascuna delle farine proteiche sostitutive. Ad esempio, lo stesso problema di sincronismo precedentemente descritto per la metionina, si viene a presentare per tutti gli AA essenziali, per esempio per la lisina (Lys), nel caso della sostituzione di FM con farina di piume, ma anche con la taurina (Tau). Quest’ultimo è un aminoacido solforato indispensabile, benché di fatto non partecipi all’architetura delle catene proteiche, è in parte biosintetizzabile nel fegato, a partire dalla cisteina, un altro aminoacido solforato derivabile dalla metionina.
Ciò ha quindi reso necessario affrontare una serie di prove preliminari, finalizzate a studiare la cronologia di assunzione e metabolizzazione degli AA addizionati, rispetto agli AA proteici. Spigole di taglia giovanile sono quindi state alimentate con mangimi commerciali a base di farina di pesce come controllo, quindi con formulazioni dove la farina di pesce era sostituita con proteine vegetali. In queste ultime si era provveduto ad un arricchimento con varie forme commerciali di metionina, col tentativo di fornire, in tutti i casi, un’adeguata inclusione di metionina. La finalità era di comparare tra di loro le varie forme aminoacidiche, sia per l’efficienza zootecnica che per la risposta metabolica. In dettaglio, sono stae utilizzate inclusioni di L-Met, DL-Met, un precursore della metionina o metionina idrossianalogo (HTMB) ed il dipeptide Met-Met.
Oltre allo studio delle performance di crescita e di conversione dell’alimento, sono stati studiati nel fegato delle spigole alcuni aspetti della risposta metabolica, studiando l’attività trascrittomica di geni chiave del ciclo della metionina, e di alcuni metaboliti intermedi.
Ciò ha quindi reso necessario affrontare una serie di prove preliminari, finalizzate a studiare la cronologia di assunzione e metabolizzazione degli AA addizionati, rispetto agli AA proteici. Spigole di taglia giovanile sono quindi state alimentate con mangimi commerciali a base di farina di pesce come controllo, quindi con formulazioni dove la farina di pesce era sostituita con proteine vegetali. In queste ultime si era provveduto ad un arricchimento con varie forme commerciali di metionina, col tentativo di fornire, in tutti i casi, un’adeguata inclusione di metionina. La finalità era di comparare tra di loro le varie forme aminoacidiche, sia per l’efficienza zootecnica che per la risposta metabolica. In dettaglio, sono stae utilizzate inclusioni di L-Met, DL-Met, un precursore della metionina o metionina idrossianalogo (HTMB) ed il dipeptide Met-Met.
Oltre allo studio delle performance di crescita e di conversione dell’alimento, sono stati studiati nel fegato delle spigole alcuni aspetti della risposta metabolica, studiando l’attività trascrittomica di geni chiave del ciclo della metionina, e di alcuni metaboliti intermedi.
I risultati relativi alla crescita hanno mostrato come atteso crescite e conversioni comparabili, caratterizzate comunque da differenze che in 3 mesi di prova non sono risultate statisticamente significative. Dai risultati dello studio in piattaforma molecolare è emerso come la forma L-Met possa essere direttamente utilizzata per la sintesi di metaboliti intermedi del ciclo della metionina, oppure essere degradata attraverso vie quali la transaminazione. Le forme D-Met, Met-Met, HTMB, sembrano presentare una disponibilità più lenta, dovendo prima essere trasformate in L-Met.
L’interpretazione dei risultati deve essere vista nel quadro dell’omeostasi dei metaboliti del ciclo della metionina, fino alla biosintesi della cisteina e della taurina. Le forme D-Met, Met-Met, HTMB, presentano presumibilmente un’assimilazione più lenta, dovendo prima essere trasformate in L-Met.
E’ comunque risultato evidente come sarebbe pertanto riduttivo limitare l’interesse verso la metionina alla sua funzione di aminoacido proteinogenico, visti i suoi molteplici e complessi ruoli nel metabolismo. Se da un lato lo studio ha mostrato in modo parzialmente inatteso che tutte le forme di metionina utilizzate sono idonee a garantire performance di crescita e conversione, tanto da apparentemente giustificarne la scelta sulla base semplicemente dei costi, d’altro lato è evidente che una disponibilità anche “anticipata” di questo aminoacido, può modificare in modo negativo alcuni equilibri nei rapporti tra importanti metaboliti del ciclo della metionina.
E’ comunque risultato evidente come sarebbe pertanto riduttivo limitare l’interesse verso la metionina alla sua funzione di aminoacido proteinogenico, visti i suoi molteplici e complessi ruoli nel metabolismo. Se da un lato lo studio ha mostrato in modo parzialmente inatteso che tutte le forme di metionina utilizzate sono idonee a garantire performance di crescita e conversione, tanto da apparentemente giustificarne la scelta sulla base semplicemente dei costi, d’altro lato è evidente che una disponibilità anche “anticipata” di questo aminoacido, può modificare in modo negativo alcuni equilibri nei rapporti tra importanti metaboliti del ciclo della metionina.
In conclusione, questa sperimentazione contribuisce alle conoscenze di base sviluppate con il progetto AGER 4F, consentendo di avviare prove di crescita in azienda con parziale o totale sostituzione della farina di pesce nei mangimi.
Fonte: Marco Saroglia, Genciana Terova e Simona Rimoldi, Università degli Studi dell'Insubria







